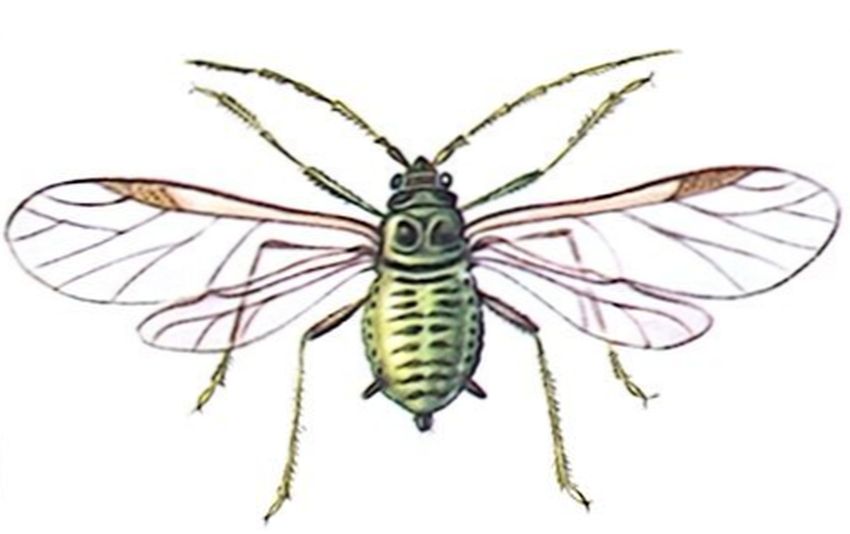
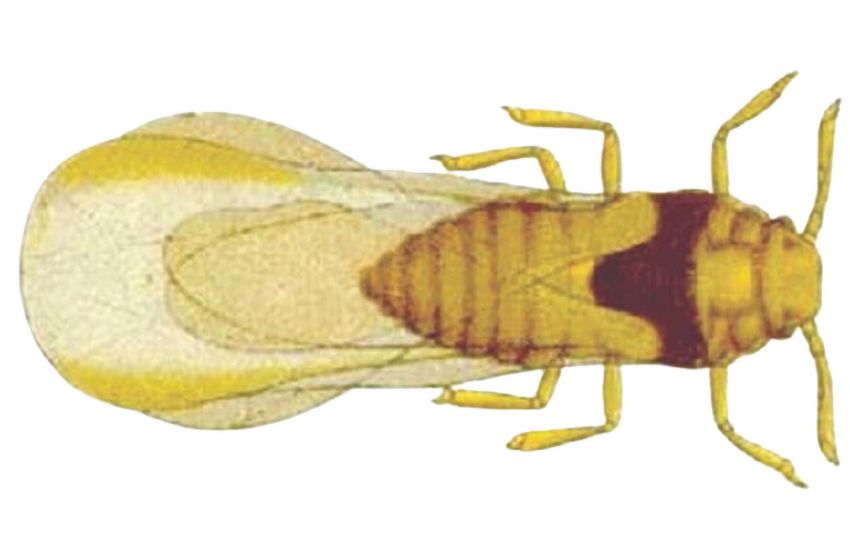
Распространенные повсеместно тли преобладают в Северном полушарии в странах умеренного климата. Они отличаются мелкими размерами (0,5—б мм), 3—6-члениковыми антеннами с острым шпилем на вершинах, наличием на крыльях (если они развиты) птеростигмы, сложными фасеточными глазами, у заднего края которых обособляется трехфасеточный бугорок, соковыми трубочками по бокам пятого сегмента брюшка, конец которого часто вытянут в виде «хвостика».
Описано около 3500 видов тлей, но каждый год обнаруживают десятки новых видов. Для многих тлей характерно чередование способов размножения (гетерогония) и поколений: обоеполого (амфигонного) и девственных партеногенетических. У тлей резко выражен полиморфизм. Нередко этому сопутствует сезонная смена растений-хозяев (гетереция) у двудомных мигрирующих (гетерецийных) видов.
У однодомных (автоцийных) видов из оплодотворенных зимующих яиц с наступлением весны вылупляются самки — основательницы колоний, которые продуцируют ряд партеногенетических поколений бескрылых и крылатых девственниц. Последние, выполняя роль расселительниц, содействуют распространению популяции на соседние растения данного вида и основывают новые колонии, где идет партеногенетическое размножение. С наступлением осени их потомки продуцируют особей-полоносок (гинопары): крылатых самцов и обычно бескрылых самок. После спаривания самки откладывают оплодотворенные яйца, которые остаются зимовать и после зимовки дают начало новым самкам- основательницам.
У двудомных видов тлей развитие происходит со сменой хозяев. При этом на первичном хозяине, обычно древесном растении, развивается амфигонное поколение и зимует оплодотворенное яйцо. Весной из него вылупляется самка-основательница, которая здесь же дает начало образованию колонии и развитию 2—3 девственных, партеногенетических поколений, в последнем из них формируются крылатые девственницы-мигранты. Перелетая на вторичного хозяина, обычно травянистое растение, эти мигранты дают начало нескольким поколениям переселенцев, но осенью формирующиеся здесь крылатые полоноски возвращаются на первичного хозяина. Таким образом, цикл замыкается, но нередко на первичном хозяине полоноски отрождают только самок, а самцы формируются на вторичном хозяине. Затем они перелетают к самкам и оплодотворяют их.
Обычно тли существуют колониями — громадными скоплениями особей, покрывающими ветви и листья растений сплошным слоем. Столь высокая плотность поселения наряду с неизбежными при этом изменениями физиологического состояния гибнущих растений служит сигналом к формированию мигрантов, расселяющихся на новые, еще не заселенные растения.
По-видимому, переход с одних растений на другие обусловлен не стремлением к расширению спектра питания, как полагали ранее, а постепенным ухудшением условий существования на истощаемом первичном хозяине. По ходу эволюционных преобразований тли переселяются с хвойных деревьев на лиственные, далее — на кустарники, а с кустарников и деревьев — на травянистые растения. При этом у некоторых видов, попавших в другие условия, упрощается жизненный цикл, например, за счет выпадения амфи- гонного поколения. Аналоциклые (то есть с неполным циклом) виды, переходящие к постоянному партеногенезу такого рода, встречаются в тропиках и субтропиках, а также в теплицах и оранжереях.
У виноградной филлоксеры — Viteus vitifolii Fitch, (греч. филлос — лист, ксеро — иссушаю), уничтожившей в конце XIX в. виноградники Франции, вместо смены растений-хозяев происходят периодические миграции с листьев виноградной лозы на ее корни. Формирующиеся здесь девственницы, выходя на поверхность, откладывают на лозе яйца, из которых развивается обоеполое поколение. После спаривания самки откладывают зимующие оплодотворенные яйца, из которых с наступлением весны выходят девственные самки, дающие начало летним поколениям, образующим галлы на листьях. Однако некоторые из них сразу уходят в почву, на корни лозы. Такой полный цикл развития наблюдается на родине вредителя, в Северной Америке. Завезенная в Европу филлоксера утратила поколения, развивающиеся на листве, и представлена исключительно корнеобитающими формами.
Полагают, что в тропики тли проникли вторично под влиянием оледенения. Косвенным доводом в пользу этого мнения служит то, что там они представлены исключительно формами с неполным циклом, относящимися к немногим семействам. Во всяком случае центром видового и биологического разнообразия подотряда следует признать зону умеренного климата.
Среди однодомных видов тли в нашей стране встречается капустная (Brevicoryne brassicae L.) и зеленая яблонная тля {Aphis роті Deg.), а среди двудомных — бобовая (свекловичная) тля (Aphis fcibae Scop.), развивающиеся сначала на бересклете, калине и чубушнике, а затем на конских бобах, свекле и других растениях.
Многие муравьи используют тлей как источник сахаристых выделений. Перенося их с одного растения на другое, муравьи содействуют расселению тлей и охраняют их от паразитов и хищников. В ограничении численности тлей важную роль играют божьи коровки, сетчатокрылые (златоглазки), личинки мух-журчалок, некоторые клопы и многие паразитические перепончатокрылые. В частности, нередкие в Подмосковье массовые размножения злаковой тли (Schizaphis gramina Rond.) обычно сопровождаются нашествиями на заселенные тлей поля семиточечных божьих коровок (Coccinella sepíempunctata L.), способных существенно ограничить наносимый вредителем ущерб урожаю.